Theory of Mind Impairment in Autism as a Result of Failed Implementation of Integrated Social Percepts
 Introduction
Introduction
Autism Spectrum Disorder (ASD) is a complex neurodevelopmental disorder characterized by an array of perceptual, social, and neurological abnormalities. Diagnoses of ASD are typically made between ages two and four and are based on broad behavioral criteria including impairments in verbal and nonverbal communication, a lack of social or emotional reciprocity, restricted repetitive and stereotyped patterns of behavior, and persistent preoccupations with parts of objects (1). Individuals with autism often may demonstrate heightened abilities in rote memory and attention to detail. These characteristics are common across different severities of the disorder. Asperger’s syndrome is a closely related, yet milder variant that is distinguished from autism by its lack of linguistic or cognitive delay.
Overall, the neuroanatomical abnormalities in autism seem to affect regions of the brain crucial for social interactions. Questions remain, however, as to how these neurological abnormalities correspond to autism’s complex array of perceptual and social deficits. Further research is also necessary to elucidate the developmental relationship between irregularities in lower-level processing areas and those in higher cognitive areas: to what extent do autism’s secondary executive functioning deficits exacerbate the neuroanatomical abnormalities in the primary perceptual areas? Current research initiatives address this question and others: is there a more quantitative means of discerning the severity and characteristics of different autism phenotypes? Can the neurological correlates of the disorder’s cognitive deficits be pinpointed?
Strategic methods employed to answer these questions include developmental studies, sibling and first-degree relative studies, lesion studies, theory of mind studies, and studies of symptomatically-related disorders (e.g., Asperger’s Syndrome, Fragile X Syndrome). Mapping the constellation of deficits and characteristics of autism will require an array of integrative and comparative studies. Certainly, these efforts will lead not only to a better understanding of an enigmatic and pervasive disorder, but also to an illumination of some of the complicated integrative processes that characterize human cognitive complexity (i.e., top-down modulation and theory of mind).
Cognitive Developmental Aspects of Autism
Autism is characterized by an array of cognitive disorders, many of which comprise the current diagnostic criteria. Some of the hallmarks include abnormal language development such as muteness, language delay, echoing of speech, and idiosyncratic use of language (2). Overall, autistic individuals display impaired verbal as well as nonverbal communication. Furthermore, autistic individuals exhibit a striking impairment in perceptual processes that require rapid attentional shifting or sensory integration (3, 4, 5). Social deficits are marked by an inability to empathize and a lack of implementation of theory of mind processes (3, 6).
Deficits in Integration of Sensory Perceptions
Many of the cognitive deficits in autism result from impairments in sensory integration (4, 5, 7). Without the ability to synthesize their perceptions, autistic individuals have difficulties interacting with their environments, particularly in social situations. A recent study by Westerfield et al. (2007) confirms previous research that crossmodal sensory integration in autism is neither early nor automatic (5). The study examined the temporal, spatial, and attentional factors that could disrupt integration of auditory and visual information. Varying the congruence of spatial and temporal proximity of simple auditory and visual stimuli revealed that autism subjects were not affected by mismatch in picture and sound except in the most extreme mismatch, in which the picture and sound were separated in time by more than 150 milliseconds. This finding suggests that auditory-visual integration takes place at a much later stage in autistic individuals than in typically-developing individuals (5). By extension, the slower integration of perceptual stimuli indicates a likely contributing factor to the social deficits characteristic of autism. The lack of rapid and automatic integration of multiple sensory percepts and social observations would impair autistic individuals in their ability to make inferences about the mental states of others and to provide context for their social interactions.
In a related study, Townsend et al. (2007) extended the hypothesis that social deficits in autism may be caused in part by impaired sensory integration. The new postulation specifies that these impairments occur in the automatic implementation of integrated perceptions. Their study employed a series of behavioral and EEG/ERP studies to examine implicit learning of a sequence rule to predict the appearance of a certain target. The authors compared the subjects in their ability to use the implicitly-learned rule to improve performance versus their ability to use the rule to improve performance when it was explicitly supplied. Results indicated that autistic individuals were not able to implement implicitly-learned information in order to improve performance, but they were able to improve their performance when the rule was made explicit. The authors suggest that autistic individuals lack an automatic implementation of sensory information, but are able to do so with conscious effort.
Normal and Impaired Theory of Mind Processes
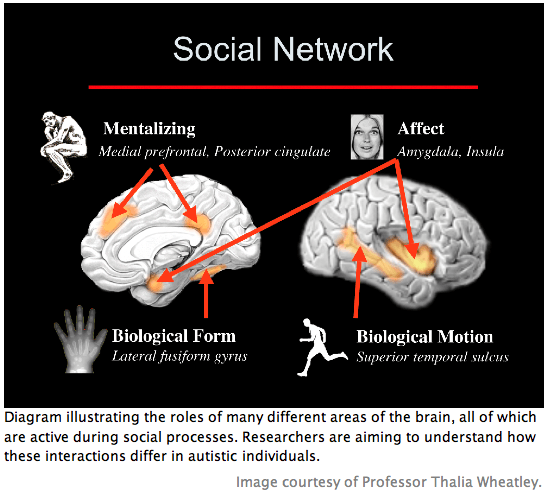
Theory of mind refers to the ability to explain and predict others’ behavior by inferring their thoughts, feelings, and motivations (2, 8). It involves rapid and automatic implementation and modulation based on context (2, 8). As far as is known, theory of mind processes are universal and follow a particular stereotyped development that is similar across individuals (2, 8). One of the earliest manifestations of a child’s mentalizing abilities is seen at around 18 months in the toddler’s enjoyment of pretence (2, 8). Playing “make believe” is a favorite game of most young children and requires an understanding of others’ mental states, including intent and belief (2). That these mentalizing abilities follow a stereotyped development suggests control by a dedicated neural mechanism (2). Structures thought to be involved include the paracingluate cortex, the temporal-parietal junction, the amygdala, the temporal poles adjacent to the amygdala, and the frontal lobes (7, 8, 9).
Most of the central deficits in autism occur in theory of mind processes. Many autistic individuals have described their bewilderment at the mysterious “mind reading” capabilities of other, normal individuals (2). Autistic children lack the ability to mentalize from birth; some of the earliest symptoms of the disorder are evident when young children fail to follow another person’s gaze or understand games of “make believe” (2). Autism’s hallmark abnormalities in language development may be a by-product of the theory of mind deficits (2). Without normal abilities to infer another’s intent or to follow another’s eye gaze, children with autism show difficulty in mapping the word to the object to which the speaker is referring.
Carrying out normal social interactions requires a constant incorporation of contextual information and prior social knowledge. This ability is impaired in autism, but it is possible for some individuals with less severe phenotypes of the disorder to carry out relatively normal social behaviors. This normal behavior, however, is accomplished by alternative compensatory processes that circumvent the persistent physiological abnormalities. High functioning autistic (HFA) individuals thus share similar atypical processing methods to individuals with more severe phenotypes (3). For example, HFA individuals display the same abnormal face gaze pattern as individuals with more severe phenotypes: aversion to direct eye gaze and increased fixation on the mouth. A recent study by Spezio et al. (2006) employed a new method of “Bubbles” to quantify the extent to which HFA subjects avoid eye gaze and instead seek information from the mouth area (10). The “Bubbles” method varies the facial information available on a given trial by revealing only small parts of the face. Measurements of the eye movements made as participants view these stimuli provide novel detail about the abnormal way in which autistic individuals look at faces. The findings by Spezio et al. showed marked differences in HFA eye gaze patterns compared to normal subjects. A possible extension of this study would be to repeat it with individuals of a more severe autistic phenotype and to compare the different patterns of face gaze. This type of analysis could lead to more specific and quantitative diagnostic criteria based on the varying extents to which individuals follow autistic face gaze patterns.
Neuroanatomical Abnormalities
Neuroanatomical abnormalities in autism are numerous and follow a complex developmental pattern. Studies of ASD are still in the process of elucidating the primary deficits. By the time a diagnosis is made, it is difficult to discern the trajectory of the neurodevelopmental abnormalities. Studies so far seem to indicate two patterns: (1) Early brain overgrowth followed by a premature cessation of growth that ultimately results in a brain size comparable to that of typically-developing individuals (but an enlarged amygdala persists); and (2) Primary deficits in lower-level processing areas that lead to aberrant development of higher cognitive areas and abnormal neural connectivity.
Early Brain Overgrowth Followed By Premature Cessation of Growth
Neuroimaging studies and head circumference data suggest that one of the earliest deviations from normal brain development involves a transient period of postnatal macrocephaly followed by a premature cessation of brain growth (11). Courchesne et al. (2001) found that maximum brain size was reached in autistic individuals by about three to five years of age, which is roughly six to 10 years earlier than typically-developing individuals (11). A recent MRI longitudinal study by Schumann et al. (2007) confirmed and extended previous studies on the pattern of brain growth in autism (12). Their work included a total of 220 clinical evaluations and MRI scans collected at roughly twelve-month intervals from 18-60 months from two groups of subjects: (1) typically-developing toddlers (n=48); and (2) toddlers with autism spectrum disorder (n=43). Each MRI underwent a detailed examination by the automated program Freesurfer, which segmented the frontal lobe into grey and white matter as well as further parcellated the cortex into superior, middle, and inferior frontal gyri, orbitofrontal cortex, frontal pole, precentral gyrus, and anterior cingulate cortex.
A preliminary regression analysis of a subset of longitudinal data revealed significant enlargement of the collective frontal lobe and frontal gray matter volumes (p<.05) in toddlers with ASD compared to typically-developing controls. These data confirm earlier findings that the frontal cortex – more specifically, the cerebrum, frontal, and temporal lobes—exhibits the highest degree of enlargement during the period of transient macrocephaly in autistic brain development (11). A possible explanation for this excessive frontal lobe volume is that the developing brain may be compensating for sensory filtering deficits in the lower-level processing areas. Without proper preliminary information processing, input to the frontal lobe is in an unusually low signal-to-noise level (i.e., the input is relatively unfiltered and nonspecific and, therefore, is difficult for the higher cognitive areas to process). This condition may lead to abnormal neural connectivity and an inability to develop normal executive functioning (11).
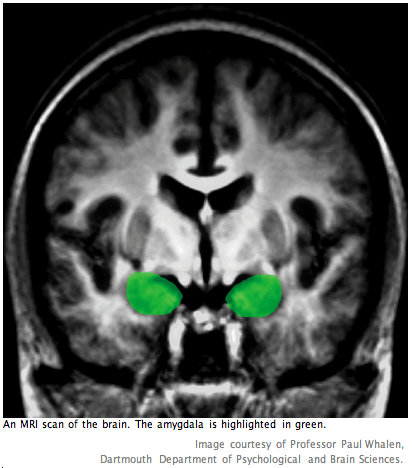
Although autistic individuals have larger brains compared to typically-developing individuals during early developmental stages, the latter catch up after the former experience a premature cessation of growth. Notably, although adults with autism possess similar brain volumes to typically-developing adults, the amygdala (which, as mentioned earlier, is thought to be a key region involved in theory of mind processes) remains enlarged (11, 13). A recent study by Nordahl and Amaral, et al. (2007) replicated and extended previous findings of increased amygdala volume in autistic individuals (13). Their work focused on investigating brain structure in very young children with autism, a relatively unstudied area of research. Nordahl et al. attribute the sparseness of MRI studies of very young children with autism to challenges in acquiring high quality data. A significant outcome of their study was the development of a new protocol for obtaining brain images of very young children. Using the fMRI to observe this age group has been notoriously difficult. Instead of resorting to sedation or anesthesia to still the children during the scan, Nordahl et al. scanned their subjects during natural, nocturnal sleep. If this new protocol is consistently successful and replicable, it may be a valuable tool for future studies that target earlier stages of brain development in autism. It is hoped that these will lead to elucidations regarding the primary neuroanatomical deficits in autism spectrum disorder.
An fMRI study by Wallace et al. (2007) confirms previous findings that autistic individuals have reduced grey matter volume and less pronounced frontal lobe asymmetry (14). Significantly, their study also extends these characteristics to correlate symptom severity with the degree of frontal lobe asymmetry. While typically-developing individuals display marked left frontal lobe asymmetry, the most impaired autistic individuals display comparably little frontal lobe asymmetry. High functioning autistic individuals (HFA) who may often achieve normal behavioral output possess less frontal lobe asymmetry than typically-developing individuals but more asymmetry than individuals with more severe autistic phenotypes. This structural data thus supports previous physiological data that normal behavioral output by HFA subjects is accomplished by abnormal compensatory neural circuits (3). The precise developmental relationship between lower-level processing areas and higher cognitive areas remains uncertain. Questions persist as to the extent to which secondary effects in higher cognitive areas exacerbate the original lower-level abnormalities. Earlier developmental studies may lead to a better understanding of the severity and characteristics of the primary neuroanatomical deficits.
Lower-Level Deficits Lead to Aberrant Development of Higher Cognitive Areas
Ultimately, autistic brains (including those of HFA individuals) show decreased grey matter in anterior regions such as the paracingulate sulcus – a region thought to be involved in theory of mind processes and, specifically, mentalizing of the self (2, 15)—and the inferior frontal gyrus (2). Conversely, increases in grey matter are found in posterior regions such as the peri-amygdaloid cortex and the middle temporal and inferior temporal gyrus (2). These findings are inconsistent with behavioral neuroimaging studies that show hyperactivation and abnormal patterning of primary perceptual processes coupled with hypoactivation of more complex cognitive processes (3). Both structural and functional data support the hypothesis that hyperarousal occurs as a result of aberrant filtering in lower-level processing regions, which leads to abnormal neural connectivity and, ultimately, impairments in top-down control.
In the typically-developing child, experience and normal neural growth processes act in tandem to develop organized and finely-tuned higher cognitive areas. Autistic individuals, however, are unable to carry out proper primary processing of their experiences. As a result, their higher cognitive areas develop abnormally in spite of intact neural growth mechanisms (7). Neuropathology studies show abnormally dense packing of neurons occurs in limbic regions such as the hippocampus and the amygdala as well as a reduction in the size of cortical minicolumns. Furthermore, cells within the minicolumns show increased dispersion. These alterations could increase the total number of minicolumns and, consequently, the degree of connectivity between minicolumns (3, 7).
The over-connectedness within cortical regions and the abnormally densely packed neurons in the limbic regions suggest that higher cognitive areas have developed in a compensatory manner in response to excessive noise in the input from the lower-level processing areas. The inhibition of higher cognitive functions in autistic individuals causes impairments in top-down modulation processes such as the selection of perceptual input1. This lack of top-down control likely contributes to deficits in theory of mind processes that require informed inferences based on an integration of real-time observations and prior experience.
 The Amygdala as a Possible Cause for Social Deficits
The Amygdala as a Possible Cause for Social Deficits
Studies of the amygdala strongly suggest that it is involved in mediating experiences of fear, anxiety, and surprise (16, 17). More specifically, the amygdala has been shown to play a role in perceiving facial emotion – particularly negative emotions such as anger—and then linking it to its social meaning (6, 15, 17, 18). Because of its important role in recognizing others’ emotions, the amygdala has become an area of interest in theory of mind studies. Furthermore, because of the volumetric enlargement of the amygdala in autistic individuals compared to typically-developing individuals, it has gained attention as a possible source of many of autism’s theory of mind deficits (2, 6, 15, 18). In order to investigate the role of the amygdala in normal and impaired theory of mind processes, studies have focused on comparing its activation in normal subjects with subjects who have amygdala. They also have focused on comparing the related behavioral output in subjects with brain damage to autistic subjects (e.g., in studies of perception of facial expression). Comparing these data may lead to a better understanding of the amygdala’s role in the neural network involved in theory of mind as well as to a more specific sense of the neural correlates of autism’s social deficits.
Some of the strongest evidence for the amygdala’s specific role in recognizing social cues from faces comes from a study how amygdala damage affects the ability to discern emotion from facial expressions (Adolphs et al., 2003) (18). Overall, bilateral amygdala damage did not seem to impair recognition of emotion from non-facial cues such as body language. All four subjects with bilateral damage, however, were significantly impaired in recognizing angry faces shown in isolation, which they frequently mistook for expressing happiness (Adolphs et al. postulate that this may be because these expressions of anger often showed bared teeth). This impairment in discerning emotion from facial expressions could lead to deficits in empathizing and in inferring the mental states of others, both of which are hallmarks of autism spectrum disorder (6).
Current challenges in the field of amygdala research are similar to those in theory of mind research: how can experimental paradigms be extended from static images to more realistic, real-time social interactions? More complicated paradigms are required to elucidate how the amygdala functions as a part of the complex neural network responsible for making subtle social inferences from an integration of real-time observations with prior knowledge and self-understanding. A recent study by Spezio et al. (2007) examined eye gaze in an individual with bilateral amygdala damage (participant S.M., a 42-year-old woman) compared to five matched controls during real-life social interactions (10). In a face-to-face session, an actor posed questions to a subject fitted with eye tracking machinery. Previous studies have shown that autistic individuals tend to avoid direct eye gaze and instead attempt to glean social information from the mouth (7, 10, 15). Significantly, Spezio et al. showed that bilateral amygdala damage in S.M. produced similar face gaze patterns to autism: there was almost no direct eye contact and nearly exclusive gaze to the mouth.
Although the study by Spezio et al. presents data from only one subject with bilateral amygdala damage, the findings are striking. These data not only confirm previous findings that autistic individuals and individuals with amygdala damage show similar cognitive deficits, but they also further indicate that these deficits seem to come from identical abnormalities in physiological processes. Together, these results strongly suggest that the amygdala is one of the central structures responsible for autism’s social deficits.
Future initiatives in amygdala and theory of mind research likely will center on elucidating how the amygdala comes into play in the neural network involved in theory of mind. A recent study by Kim et al. (2004) examined the differential responses of the amygdala to surprised faces based on varying contextual information (17). Normal, healthy subjects viewed faces showing surprised expressions, which were preceded by either a positive sentence or a negative sentence. Using fMRI neuroimaging, Kim et al. examined the functional relationship between the amygdala and the medial prefrontal cortex with regard to their differential responses based on the context given to the surprised facial expression by the preceding sentence. Results indicated that responses to negative versus positive sentences were greater within the ventrolateral prefrontal cortex, while responses to positive versus negative sentences were greater within the ventromedial prefrontal cortex (17). But what is the machinery responsible for this connectivity between the amygdala and the medial prefrontal cortex? Is there a third brain region involved, or are morphological constructs such as glia responsible for this critical networking? Uncovering the interconnections of the neural network involved in theory of mind processes will lead to a better understanding of autism’s social deficits and to a clearer sense of the neural mechanisms that govern human social interactions.
Current and Future Directions
In the near future, autism and theory of mind studies can begin to look toward illuminating a more complicated neural network. The priority of most current research initiatives, however, remains to elucidate the structures and behaviors involved in isolation. There is still much to learn about the specific behavioral functions of many neurological structures and the details of autism’s abnormal neurodevelopmental trajectory.
One major area of study compares the cognitive behaviors and physiological characteristics of autistic individuals to those of their first-degree relatives. A recent fMRI study by Belmonte et al. (2007) examined the physiology of visual attention in autism families (19). Their results showed that both autistic individuals and their non-autistic siblings were impaired at a visual divided-attention task and displayed atypical frontal activation in comparison to normal controls. Analysis of correlations across brain regions, however, revealed that autistic individuals exhibited decreased functional correlation compared to normal siblings. Belmonte et al. thus suggest that although atypical frontal activation may reflect processes that are permissive but not determinative of autistic brain development. Future research likely will attempt to illuminate these determinative factors, which may be related to functional interconnectedness across an array of brain regions.
Other current research initiatives are exploring symptomatically-related disorders. Asperger’s Syndrome is a disorder that often is regarded as a “less severe” form of autism and a more manageable venue in which to study cognitive and social deficits. An fMRI study by Moran et al. (2007) investigated self-referential processing, a process integral to theory of mind functions in individuals with Asperger’s Syndrome (AS) (20). When asked to attribute adjectives to themselves and to others, normal subjects did so significantly faster and with more robust activation in cortical midline structures such as the medial prefrontal cortex and the posterior cingulate cortex compared to AS subjects.
It is notable to recall that findings by Kim et al. (2004) suggest that the medial prefrontal cortex is connected to the amygdala in a network involved in contextual processing of emotion from facial expressions. Therefore, certain theory of mind deficits in inferring the emotional states of others – specifically from facial expressions—may come from impairments in this collaborative process by the medial prefrontal cortex and the amygdala. In autism, an enlarged amygdala likely causes excessive activation of fear and anxiety responses. A weakly activated medial prefrontal cortex may lead to an impaired ability to empathize (i.e., to use self-referential knowledge of one’s own emotions), which could inhibit the contextual modulation of the amygdala’s response, thereby further magnifying the amygdala’s fear and anxiety response. Such a combined effect could help to explain how autistic individuals might develop an abnormal aversion to direct eye gaze.
Advances in neuroimaging may help to obtain earlier and higher quality data in developmental studies. Increased detail in images taken during sensory perception and theory of mind tasks could reveal more specific information about which regions are being activated and how they interconnect. Furthermore, developmental studies may explore morphological abnormalities in children with autism. A recent study by Morgan et al. (2007) suggests that examining the development of glial infrastructure could lead to important information about the developmental abnormalities in autism (21). The study examined microglial cell populations in a 3.8-year-old with autism and a 1.8-year-old control. Results indicated that robust activation of microglial cells – normally seen after there has been trauma to the brain—can occur in the autistic brain at an early developmental age. Although the subject pool was limited, the data obtained recommends further investigation of microglia development in larger samples of young autistic cases. Given that glia is emerging in the larger field of neuroscience as an increasingly important neurobiological construct, it may be a valuable area of study in autism research. The widespread interconnectivity of glia in the brain suggests that glial abnormalities may contribute to the atypical functional connectivity found in autism.
Future research using lesion studies and more advanced and realistic theory of mind paradigms will lead to a better understanding of autism’s cognitive deficits. Experimental tasks that more closely resemble real-life social interactions could provide more detailed information about the neural and physiological mechanisms involved in complex social behavior. Comparing the functioning of these mechanisms in control subjects to autistic subjects could help to reveal the locus (or loci) of autistic impairments in implementing integrated perceptual information and prior knowledge (22). Particular challenges in creating these new paradigms will involve segregating which elements are testing theory of mind and which are testing other aspects of social interactions such as rapid attentional shifting. Isolating regions of interest will also be an obstacle in lesion studies, in which finding subjects with specific patterns of brain damage can be exceedingly difficult (3).
A possible starting point in lesion studies may be to continue investigating eye gaze and recognition of emotion from facial expressions. One study of interest might be to compare amygdala activation in autistic individuals versus controls during direct eye gaze. Given that autistic individuals have an enlarged amygdala and seem to show aversion to direct eye gaze, the expectation would be to see hyperactivity in the autistic amygdala compared to controls. Another study might repeat the experiment developed by Kim et al. (2004), except using subjects with brain damage to the medial prefrontal cortex. Comparing the performance of brain damaged subjects to that of control subjects might illuminate more about the role of the medial prefrontal cortex in the contextual modification of amygdala responsivity. Because amygdala studies most often involve negatively-valenced emotions, a difficulty in any amygdala study will be to keep anxiety levels low across subjects. This will likely be particularly challenging in autistic subjects, for whom direct face gaze and social interactions are unusually anxiety-provoking. Steps to alleviate this anxiety might involve providing substantial practice training for all subjects as well as frequent rest periods in between trials (3).
A priority in current and future autism studies will be to delineate more specific diagnostic criteria. Quantitative gradations of the autism phenotype might be developed as more knowledge is gained about the determinative characteristics of the disorder. A significant study by Chiu et al. (2007) has developed a preliminary algorithm for determining the severity of different autism phenotypes (4). This algorithm is called a “self-eigenvalue” and was derived from fMRI studies of cingulate cortex activation during self-referential studies. Primary evidence for the algorithm was provided by a related study by Kishida et al. (2007), which found striking correlations in cingulate activation in the brains of Division I athletes (23). Neuroimaging data indicated an inverse activation of the cingulate cortex when athletes viewed clips of other athletes playing sports of their expertise compared to when they were subsequently asked to “do it” (i.e., visualize themselves carrying out the action in the clip). Chiu et al. observed a similar pattern of cingulate activation in control subjects during an active social exchange game with a human partner (the activation disappeared in the absence of an interactive social partner). Significantly, high functioning autistic subjects lacked the cingulate response pattern of control subjects. Perhaps most striking is that this activation was not “all-or-none,” but rather the extent of cingulate activation varied parametrically with symptom severity. An algorithm developed from the parametric relation between cingulate activation and symptom severity would be an important step in outlining quantitative autism phenotypes.
Conclusion
Progress in autism research is contingent upon expanding our understanding of complex cognitive processes such as top-down modulation and theory of mind. Future studies will require new and complex paradigms that come closer to recreating the complex social interactions in which these neural networks are activated. For the present, primary goals in autism research include gaining a better understanding of the complex pattern of deficits in autism and developing a more precise means of diagnosing different autistic phenotypes. Hopefully, these advances will eventually lead to finding a means of helping even severely autistic individuals to reach a higher level of social functioning.
References
1. American Psychiatric Association, Diagnostic and Statistical Manual of Mental Disorders: Fourth Edition. (American Psychiatric Pub., Inc., 2000). Available at http://www.autism-watch.org/general/dsm.shtml
2. U. Frith, Neuron 32, 969 (2001).
3. M. Belmonte and D. Yurgelun-Todd, Cognitive Brain Research (17)3, 651 (2003).
4. P. Chiu, P. Montague, et al., Active interpersonal exchange evokes quantitative neural phenotype for high functioning autism. Program No. 304.19. 2007 Neuroscience Meeting Planner. San Diego, CA: Society for Neuroscience, 2007. Online.
5. M.A. Westerfield , P.T. Lai, G.L. Smith, A. Lincoln, J. Townsend, Auditory-Visual Integration in Autism. Program No. 175.15. 2007 Neuroscience Meeting Planner. San Diego, CA: Society for Neuroscience, 2007. Online.
6. V.E. Stone, S. Baron-Cohen, A. Calder, et al., Neuropsychologia 41, 209 (2003).
7. M. K. Belmonte, E.H. Cook, Jr., G.M. Anderson, J.L.R. Rubenstein, W.T. Greenough, A. Beckel-Mitchener, E. Courchesne, E., L.M. Boulanger, et al., Molecular Psychiatry 9, 646 (2004).
8. V.E. Stone, S. Baron-Cohen, R.T. Knight, Journal of Cognitive Neuroscience 10(5), 640 (1998).
9. K.K.W. Kampe, C.D. Frith, U. Frith, The Journal of Neuroscience 23(12), 5258 (2003).
10. M.L. Spezio, R. Adolphs, R.S.E. Hurley, J. Piven, Neuropsychologia 45, 144 (2007).
11. E. Courchesne, Mental Retardation and Developmental Disabilities Research Reviews 10, 106 (2004).
12. C. Schumann, G.M. Wideman, C. Carter Barnes, J.A. Buckwalter, D.J. Hagler, Jr., R.A. Carper, E. Courchesne, MRI longitudinal study of the frontal cortex through early childhood in autism. Program No. 61.5. 2007 Neuroscience Meeting Planner. San Diego, CA: Society for Neuroscience, 2007. Online.
13. C.W. Nordahl, T.J. Simon, and D.G. Amaral, Amygdala Enlargement in Very Young Children with Autism. Program No. 61.7. 2007 Neuroscience Meeting Planner. San Diego, CA: Society for Neuroscience, 2007. Online.
14. G.L. Wallace, J. Silvers, A. Martin, and J. Gledd, Reduced Gray Matter Volume and Frontal Lobe Asymmetry in High Functioning Autism Spectrum Disorders. Program No. 172.9. 2007 Neuroscience Meeting Planner. San Diego, CA: Society for Neuroscience, 2007. Online.
15. M.L. Spezio, S. Po-Yin Huang, F. Castelli, and R. Adophs, The Journal of Neuroscience 27(15), 3994 (2007).
16. M. Davis, and P.J. Whalen, Molecular Psychiatry 6, 13 (2001).
17. H. Kim, L.H. Somerville, T. Johnstone, S. Polis, A.L. Alexander, L.M. Shin, and P.J. Whalen, Journal of Cognitive Neuroscience 16(10), 1730 (2004).
18. R. Adolphs, D. Tranel, Neuropsychologia 41, 1281 (2003).
19. M.K. Belmonte, M. Gomot, S. Baron-Cohen, Visual Attention in Autism Families: ‘Unaffected’ Sibs Share Atypical Frontal Activation But Not Atypical Functional Connectivity. Program No. 61.14. 2007 Neuroscience Meeting Planner. San Diego, CA: Society for Neuroscience, 2007. Online.
20. J.M. Moran, A. Qureshi, M. Singh, J.D.E. Gabrieli, Neural Underpinnings of Self-Referential Processing in Asperger’s Syndrome. Program No. 304.18. 2007 Neuroscience Meeting Planner. San Diego, CA: Society for Neuroscience, 2007. Online.
21. J.T. Morgan, G. Chana, J. Buckwalter, E. Courchesne, I.P. Everall, Microglial Activation in a 3-Year-Old Autistic Brain. Program No. 172.12. 2007 Neuroscience Meeting Planner. San Diego, CA: Society for Neuroscience, 2007. Online.
22. M. Belmonte and D. Yurgelun-Todd, Cognitive Brain Research (17)3, 651 (2003).
23. K. Kishida, P. Chiu, R. Montague et al., “Self” responses revealed independently by fMRI during visualization. Program No. 304.16. 2007 Neuroscience Meeting Planner. San Diego, CA: Society for Neuroscience, 2007. Online.
Leave a Reply